Keratin (), sıklıkla skleroproteinler olarak adlandırılan yapısal lifli proteinler ailesine aittir. Bu protein, tetrapod omurgalılarında pulların, saçların, tırnakların, tüylerin, boynuzların, pençelerin, toynakların ve epidermal tabakanın birincil yapısal bileşeni olarak hizmet eder. Ayrıca keratin, epitel hücrelerine hasara veya mekanik strese karşı koruma sağlar. Özellikle keratin hem sulu hem de organik çözücülerde olağanüstü çözünmezlik sergiler. Bireysel keratin monomerleri demetler halinde polimerleşerek sürüngenler, kuşlar, amfibiler ve memelilerde gözlenen sağlam, mineralize olmayan epidermal uzantıları oluşturan ara filamentler oluşturur. Aşırı keratinizasyon, sığırların ve gergedanların boynuzları ve armadilloların osteodermleri gibi belirli dokuların yapısal olarak güçlendirilmesine katkıda bulunur. Kitin, keratinize dokularla kıyaslanabilir dayanıklılığıyla tanınan diğer tek biyolojik maddedir. Keratin iki ana formda bulunur: tüm omurgalılarda bulunan ilkel, daha yumuşak bir varyant ve yalnızca sauropsidlerde (sürüngenler ve kuşlar) bulunan daha sert, türetilmiş bir varyant.
Keratin (), skleroproteinler olarak da bilinen yapısal lifli proteinler ailesinden biridir. Dört ayaklı omurgalılarda pulları, saçları, tırnakları, tüyleri, boynuzları, pençeleri, toynakları ve derinin dış katmanını oluşturan temel yapısal malzemedir. Keratin ayrıca epitel hücrelerini hasara veya strese karşı korur. Keratin suda ve organik çözücülerde son derece çözünmez. Keratin monomerleri, sert olan ve sürüngenlerde, kuşlarda, amfibilerde ve memelilerde bulunan güçlü, mineralize olmayan epidermal uzantıları oluşturan ara filamentleri oluşturmak üzere demetler halinde birleşir. Aşırı keratinizasyon, sığır ve gergedan boynuzları ve armadilloların osteodermi gibi belirli dokuların güçlendirilmesine katkıda bulunur. Keratinleşmiş dokunun dayanıklılığına yakın olduğu bilinen diğer tek biyolojik madde kitindir. Keratin iki türde gelir: Tüm omurgalılarda bulunan ilkel, daha yumuşak formlar ve yalnızca sauropsidler (sürüngenler ve kuşlar) arasında bulunan daha sert, türetilmiş formlar.
Oluşum Örnekleri
Alfa-keratinler (α-keratinler) tüm omurgalı türlerinde her yerde bulunur. Bu proteinler memelilerin saçını (yün dahil), epidermal tabakasını, boynuzlarını, tırnaklarını, pençelerini ve toynaklarını ve ayrıca hagfish'in ürettiği sümük ipliklerini oluşturur. Ek olarak, filtreyle beslenen deniz memelilerinin balenli plakaları keratinden oluşur. Keratin filamanları, epidermisin kornifiye tabakasındaki keratinositler içinde oldukça konsantredir ve keratinizasyon sürecinden geçmiş proteinleri temsil eder. Ayrıca genellikle çeşitli epitel hücre tiplerinde bulunurlar. Örneğin, fare timik epitel hücreleri, keratin 5, keratin 8 ve keratin 14'e özgü antikorlarla reaktivite sergiler. Bu tür antikorlar, timusun genetik araştırmalarında fare timik epitel hücrelerinin alt gruplarını ayırt etmek için floresan belirteçler olarak görev yapar.
Daha sert beta-keratinler (β-keratinler), mevcut tüm sürüngenleri ve kuşları kapsayan sauropsidlerde özel olarak bulunur. Bunlar sürüngen tırnaklarının, pullarının ve pençelerinin, bazı sürüngen kabuklarının (Testudines) ve kuş tüylerinin, gagalarının ve pençelerinin bileşenleridir. Yapısal olarak bu keratinler ağırlıklı olarak beta tabakalar halinde düzenlenmiştir. Bununla birlikte, α-keratinlerin içinde beta tabakaları da gözlenmektedir. Çağdaş araştırmalar, sauropsid β-keratinler ve α-keratinler arasında temel bir genetik ve yapısal farklılığa işaret etmektedir. Sonuç olarak, yeni terminoloji korneus beta proteini'nin (CBP), α-keratinlerle ilgili belirsizliği önlemek için önerilmiştir.
Sitokeratinler olarak da adlandırılan keratinler, omurgalılar, amfioksi ve ürokordatlar da dahil olmak üzere yalnızca kordatlarda tanımlanan polimerik tip I ve tip II ara filamanlardır. Buna karşılık, nematodlar ve kordalı olmayan diğer birçok organizmanın, yalnızca nükleer yapısal lifler olarak işlev gören V tipi ara filamentlere sahip olduğu görülüyor.
Genetik Temel
İnsan genomu, 12. ve 17. kromozomlarda iki farklı küme halinde düzenlenmiş 54 fonksiyonel keratin genini kapsar. Bu genomik düzenleme, bu spesifik kromozomlar üzerindeki bir dizi gen kopyalanması olayı yoluyla evrimsel kökenlerini ima eder.
Keratin ailesi, KRT23, KRT24, KRT25, KRT26, KRT27, KRT28, KRT31, KRT32, KRT33A, KRT33B, KRT34, KRT35, KRT36, KRT37, KRT38, KRT39, KRT40, KRT71, KRT72, KRT73 dahil olmak üzere çok sayıda protein içerir. KRT74, KRT75, KRT76, KRT77, KRT78, KRT79, KRT8, KRT80, KRT81, KRT82, KRT83, KRT84, KRT85 ve KRT86, tanımlanan ilk 20 türün ötesinde keratinleri karakterize etmek için kullanılmıştır.
Yapısal Özellikler
Keratinlerin ilk dizilimi, 1982 ve 1983'te Israel Hanukoğlu ve Elaine Fuchs tarafından gerçekleştirildi. Bu diziler, daha sonra tip I ve tip II keratinler olarak adlandırılan iki farklı ancak homolog keratin ailesinin varlığını aydınlattı. Hanukoğlu ve Fuchs, bu keratinlerin ve diğer ara filaman proteinlerinin birincil yapılarının analizi yoluyla, keratinlerin ve ara filaman proteinlerinin, beta-dönüşü konformasyonunu benimsediği tahmin edilen üç kısa bağlayıcı segmentin serpiştirdiği dört a-helisel segment içeren, yaklaşık 310 kalıntıdan oluşan bir merkezi alana sahip olduğu yapısal bir model önerdiler. Keratin sarmal alanının kristal yapısının daha sonra belirlenmesi bu modeli doğruladı.
Sınıflandırma: Tip I ve Tip II Keratinler
İnsan genomu, 28'i tip I keratin ve 26'sı tip II keratinden oluşan, işlevsel olarak açıklamalı 54 keratin geni içerir.
Fibröz keratin molekülleri, son derece stabil, solak bir süperhelikal motif oluşturmak için süpersarılmaya tabi tutulur ve bu, daha sonra çok sayıda keratin monomer kopyasından oluşan filamentler halinde multimerizasyonu kolaylaştırır.
Kıvrımlı-sarmal yapısını stabilize eden birincil kuvvet, keratinin sarmal bölümleri boyunca yer alan apolar kalıntılar arasında meydana gelen hidrofobik etkileşimlere atfedilir.
Kısıtlı iç hacim, cilt, kıkırdak ve kemikte yaygın olarak bulunan, ilgisiz bir yapısal protein olan kolajenin üçlü sarmalında gözlenen yüksek glisin içeriğini belirler. Benzer şekilde, bağ dokusu proteini elastini önemli oranda hem glisin hem de alanin içerir. Yapısal proteinler tipik olarak küçük, reaktif olmayan yan zincirlere sahip amino asitlerin baskınlığıyla karakterize edilir; burada hidrojen bağlı sıkı paketleme, belirli kimyasal etkileşimlere göre önceliklidir.
Disülfür Köprüleri
Molekül içi ve moleküller arası hidrojen bağlarının ötesinde, keratinlerin tanımlayıcı bir özelliği, kükürt içeren bir amino asit olan sistein içeriğinin bol olmasıdır. Sistein, protein olmayan kükürt köprülerinin vulkanize kauçuğu stabilize etme şekline benzer şekilde, kalıcı, termal olarak stabil çapraz bağlanma yoluyla arttırılmış güç ve sertlik kazandıran disülfür köprülerinin oluşturulması için gereklidir. Örneğin insan saçı yaklaşık %14 oranında sistein içerir. Saç ve cildin yanmasıyla ortaya çıkan belirgin keskin kokular, uçucu kükürt bileşiklerinin oluşmasından kaynaklanır. Disülfür bağının kapsamlı doğası, yalnızca belirli ayrıştırıcı veya indirgeyici maddelerdeki istisnalar dışında, keratinleri büyük ölçüde çözünmez hale getirir.
Daha fazla esneklik ve esneklik sergileyen saç keratinleri, memelilerin tırnaklarında, toynaklarında ve pençelerinde bulunan keratinlerle karşılaştırıldığında daha az zincirler arası disülfit köprüsüne sahiptir; bunlar homolog, daha sert ve diğer omurgalı sınıflarındaki benzerlerine daha benzer yapılardır. Saçtakiler de dahil olmak üzere alfa-keratinler, a-helisler halinde sarılmış (düzenli zincir içi hidrojen bağına sahip) tekli protein iplikçiklerinden oluşur ve bunlar daha sonra daha fazla sarılmaya maruz kalabilecek süper sarmal halatlara bükülür. Tersine, sürüngenlerin ve kuşların karakteristiği olan β-keratinler, daha sonra disülfit köprüleri ile stabilize edilip sertleştirilen bükülmüş β-kıvrımlı tabakalardan oluşur.
Tiyollenmiş polimerler veya tiomerler, keratinlerin sistein alt yapılarıyla disülfit köprüleri oluşturabilir, böylece bu proteinlere kovalent bağlar kurabilirler. Sonuç olarak tiomerler saçta, ciltte ve çok sayıda hücre tipinin yüzeyinde bulunan keratinlere önemli derecede bağlanma afinitesi gösterir.
Filament Oluşumu
Tarihsel olarak keratinlerin 'sert' ve 'yumuşak' formlara veya alternatif olarak 'sitokeratinler' ve 'diğer keratinler'e sınıflandırılması önerildi. Bu sınıflandırma modelinin artık doğru olduğu kabul edilmektedir. 2006 yılında tanıtılan revize edilmiş bir terminoloji, bu anlayışı keratin tanımına dahil ediyor.
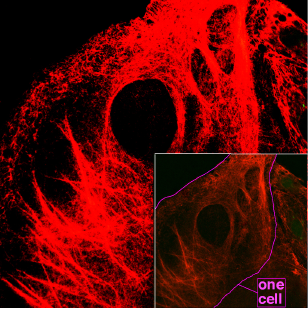
Keratin filamentleri, ara filamentler sınıfına aittir. Diğer ara filamentlerle tutarlı olarak keratin proteinleri, filamentli polimerler oluşturmak için sıralı bir birleşme sürecinden geçer. Bu süreç dimerizasyonla başlar, ardından dimerlerin tetramerlere ve ardından oktamerlere toplanması gelir. Sonuçta, geçerli hipoteze göre, bu yapılar, uzatılmış filamentler oluşturmak için uçtan uca tavlanabilen birim uzunluktaki filamentler (ULF'ler) halinde birleşir.
Eşleştirme
Kornifikasyon
Kornifikasyon, tabakalı skuamöz epitel dokusu içinde epidermal bir bariyer oluşturmaktan sorumlu olan biyolojik süreci belirtir. Hücresel ölçekte bu süreç şu şekilde karakterize edilir:
- keratin sentezi;
- plazma zarının altında yer alan kornifiye bir hücre zarfının oluşumuna toplu olarak katkıda bulunan küçük prolince zengin (SPRR) proteinlerin ve transglutaminazın üretilmesi;
- terminal hücresel farklılaşma;
- ve kornifikasyonun ileri aşamaları sırasında çekirdeklerin ve sitoplazmik organellerin nihai kaybı.
Hücresel metabolizma durur ve hücrelerin neredeyse tamamı keratin tarafından işgal edilir. Epitelyal farklılaşma boyunca, keratin proteini uzun keratin ara filamanlarına entegre olurken hücreler kornifikasyona uğrar. Sonuçta çekirdek ve sitoplazmik organeller bozulur, metabolik aktivite durur ve tam keratinizasyona ulaşıldığında hücreler programlı ölüme uğrar. Buna karşılık, dermal hücreler gibi çok sayıda başka hücre tipinde, keratin filamentleri ve diğer ara filamentler, hücre iskeletinin ayrılmaz bileşenleri olarak görev yapar ve fiziksel stres etkenlerine karşı mekanik stabilite sağlar. Bu stabilizasyon, hücre-hücre bağlantı plakları olan desmozomlara ve hücre tabanı zarı yapışkan yapıları olan hemidesmozomlara bağlantılar yoluyla elde edilir.
Epidermal hücreler, bu en dıştaki kutanöz tabakayı suya karşı büyük ölçüde geçirimsiz hale getiren ve kolajen ve elastin ile birlikte cilde gerilme mukavemeti kazandıran keratinden oluşan yapısal bir matrise sahiptir. Sürtünme ve basınç gibi mekanik uyarılar, dıştaki kornifiye epidermal tabakanın kalınlaşmasına neden olarak koruyucu nasır oluşumuna yol açar. Bu nasırlar özellikle sporcular ve yaylı çalgı çalan müzisyenlerin parmak uçları için faydalıdır. Keratinize epidermal hücreler sürekli olarak pul pul dökülür ve ardından yenilenir.
Bu sağlam bütünsel yapılar, özel dermal matrisler tarafından üretilen ölü, kornifiye hücrelerden türetilen liflerin hücreler arası çimentolaşmasından kaynaklanır. Saç sürekli bir büyüme sergilerken tüyler erime ve yenilenme döngülerinden geçer. Bileşen proteinler filogenetik homolojiyi paylaşsa da, kimyasal bileşimlerinde ve moleküller üstü mimarilerinde farklılıklar gösterirler. Karmaşık evrimsel bağlantılar yalnızca kısmen aydınlatılmıştır. Tüylerde β-keratinleri kodlayan çok sayıda gen tespit edilmiştir; bu özellik muhtemelen tüm keratin türlerine uygulanabilir.
Klinik Önem
Anormal keratin büyümesi, keratoz, hiperkeratoz ve keratoderma gibi çeşitli patolojik durumlarda kendini gösterir.
Keratin gen ekspresyonunu etkileyen mutasyonlar, aşağıdakiler de dahil olmak üzere çeşitli bozuklukların etiyolojisinde rol oynar:
- Alopesi Areata
- Epidermolizis bülloza simpleks
- Siemens'in iktiyoz bülloza
- Epidermolitik hiperkeratoz
- Steatositoma multipleks
- Keratoz farenjis
- Rabdoid fenotipli büyük hücreli akciğer karsinomunda rabdoid hücre oluşumu
Tinea pedis (atlet ayağı) ve tinea corporis (saçkıran) dahil çok sayıda hastalık, keratini metabolize eden bulaşıcı mantarlardan kaynaklanır.
Keratin, yutulduğunda sindirim asitlerine karşı önemli bir direnç gösterir. Kedilerin bakım davranışları rutin olarak tüylerin yutulmasını içerir ve bu durum, daha sonra kusulan veya dışkı yoluyla ortadan kaldırılan trikobezoarların (tüy yumakları) ilerleyici oluşumuyla sonuçlanabilir. İnsan deneklerde trikofaji, son derece nadir fakat potansiyel olarak öldürücü bir mide-bağırsak bozukluğu olan Rapunzel sendromu riskini taşır.
Teşhis Uygulamaları
Keratin ekspresyonunun analizi, anaplastik malignitelerin epitelyal kökeninin belirlenmesinde önemli ölçüde yardımcı olur. Keratin ekspresyonu sergileyen neoplazmalar karsinomları, timomaları, sarkomları ve trofoblastik tümörleri kapsar. Ayrıca keratin alt tiplerinin spesifik ekspresyon profili, metastatik lezyonların değerlendirilmesi sırasında primer tümörün kökeninin tahmin edilmesini kolaylaştırır. Örneğin, hepatoselüler karsinomlar karakteristik olarak CK8 ve CK18'i eksprese eder ve kolanjiyokarsinomlar CK7, CK8 ve CK18'i sergilerken metastatik kolorektal karsinomlar tipik olarak CK20'yi eksprese eder ancak CK7'den yoksundur.
Keratinle ilişkili proteinler (KRTAP'ler)
- Keratinle ilişkili proteinler (KRTAP'ler)
- Keratinlerdeki mutasyonların neden olduğu cilt rahatsızlıklarının listesi
- İnsanın kabuk sisteminde ifade edilen keratinlerin listesi
- Keratinlerin listesi
- Keratinaz
Referanslar
- Hair-Science.com'un saçın mikroskobik unsurları hakkındaki yazısı
- Keratinlerle ilgili Proteopedia sayfası