Les ampoules de Lorenzini (singulier : ampoule) fonctionnent comme des électrorécepteurs, des organes sensoriels spécialisés capables de détecter des champs électriques. Ces structures se manifestent par un réseau de pores remplis de mucus incrustés dans la couche cutanée des poissons cartilagineux, notamment les requins, les raies et les chimères, ainsi que chez certains poissons osseux basaux tels que le roseau, l'esturgeon et le poumon. Leur origine évolutive est liée aux organes mécanosensoriels de la ligne latérale caractéristiques des premiers vertébrés. Notamment, la majorité des poissons osseux existants et tous les vertébrés terrestres ont subi la perte de ces ampoules.
Ampoules de Lorenzini (sg. : ampoule) sont des électrorécepteurs, des organes sensoriels capables de détecter des champs électriques. Ils forment un réseau de pores remplis de mucus dans la peau des poissons cartilagineux (requins, raies et chimères) et des poissons osseux basaux tels que le roseau, l'esturgeon et le poumon. Ils sont associés et ont évolué à partir des organes mécanosensoriels de la ligne latérale des premiers vertébrés. La plupart des poissons osseux et des vertébrés terrestres ont perdu leurs ampoules de Lorenzini.
Contexte historique
Les ampoules ont été documentées pour la première fois par Marcello Malpighi, avec une description anatomique précise fournie ensuite par le médecin et ichtyologiste italien Stefano Lorenzini en 1679, bien que leur fonction physiologique reste à l'époque inélucidée. Au cours du 20ème siècle, les investigations électrophysiologiques ont indiqué une réactivité aux variations thermiques, aux stimuli mécaniques et potentiellement à la salinité. Une découverte cruciale a eu lieu en 1960, lorsque les ampoules ont été définitivement identifiées comme organes récepteurs spécialisés pour la détection des champs électriques. De plus, les premières recherches sur les canaux potassiques activés par le calcium faisaient fréquemment référence à des études menées sur l'ampoule de Lorenzini dans les patins.
Développement évolutif
Les ampoules de Lorenzini présentent une relation physique et évolutive avec les organes mécanosensoriels de la ligne latérale trouvés chez les vertébrés ancestraux. L'électroréception passive, médiée par ces ampoules, est considérée comme une caractéristique ancestrale des vertébrés, impliquant sa présence chez leur dernier ancêtre commun. Ces structures sont observées chez les poissons cartilagineux (requins, raies et chimères), les poumons, les bichirs, les cœlacanthes, les esturgeons, les spatules, les salamandres aquatiques et les céciliens. Alors que l'on pense que les ampoules de Lorenzini ont été perdues au début des trajectoires évolutives des poissons osseux et des tétrapodes, les preuves existantes concernant leur absence dans de nombreux taxons restent partielles et peu concluantes.
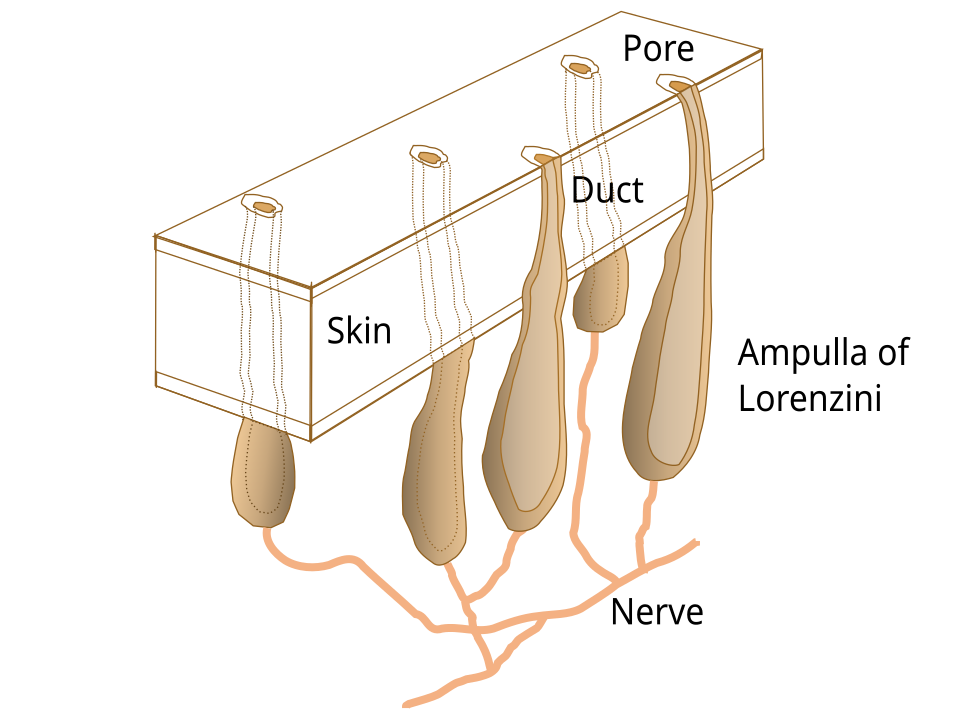
Structure anatomique
Chaque ampoule comprend un groupe de cellules sensorielles, abritant plusieurs fibres nerveuses dans un bulbe sensoriel (appelé endampulle), entouré d'une gaine de collagène. Cet ensemble se connecte à un canal rempli de gel (l'ampullengang) qui se termine au niveau d'un pore dermique à la surface de l'organisme. Le gel lui-même est une matrice à base de glycoprotéines, présentant une résistivité équivalente à celle de l'eau de mer et possédant des caractéristiques électriques analogues à celles d'un semi-conducteur.
Les pores sont principalement localisés dans les régions dermiques entourant le museau et la cavité buccale des requins et des raies, s'étendant jusqu'à des structures telles que le lambeau nasal antérieur, les barbillons, le pli circumnarial et le sillon labial inférieur. Bien que les dimensions des canaux soient généralement adaptées à la taille globale du corps de l'animal, le nombre total d'ampoules reste cohérent. Les canaux des ampoules de Lorenzini peuvent être classés comme étant poreux ou non poreux. Les canaux non poreux ne s'engagent pas dans la dynamique des fluides externes ; au lieu de cela, ils fonctionnent comme des récepteurs tactiles, atténuant les interférences des particules étrangères.
Mécanisme électroréceptif
Les ampoules sont chargées de détecter les champs électriques ambiants dans les environnements aquatiques, en détectant spécifiquement la différence de potentiel entre la tension présente au niveau des pores dermiques et la tension au niveau de la région basale des cellules électroréceptrices.
Un stimulus positif appliqué aux pores entraîne une réduction de la fréquence de l'influx nerveux provenant des cellules électroréceptrices, tandis qu'un stimulus négatif des pores provoque une augmentation de cette fréquence. Chaque ampoule est composée d'une strate singulière de cellules réceptrices, entrecoupées de cellules de soutien. Ces cellules sont interconnectées par des jonctions apicales serrées, empêchant efficacement les fuites de courant paracellulaire. Les membranes apicales des cellules réceptrices sont caractérisées par une surface limitée et une densité élevée de canaux calciques dépendants de la tension, qui déclenchent la dépolarisation, aux côtés de canaux potassiques activés par le calcium, qui facilitent la repolarisation ultérieure.
En raison de la résistance électrique exceptionnellement élevée de la paroi du canal, la totalité du différentiel de tension entre les pores du canal et l'ampoule est dissipée à travers l'épithélium récepteur d'environ 50 microns d'épaisseur. Étant donné la résistance comparativement plus faible des membranes basales des cellules réceptrices, la majorité de cette chute de tension se produit sur les faces apicales excitables, qui sont maintenues à leur seuil d'activation. Un afflux d'ions calcium à travers les cellules réceptrices induit une dépolarisation des faces basales, aboutissant à un potentiel d'action substantiel - une onde de dépolarisation se propageant suivie d'une repolarisation, analogue à celle observée dans une fibre nerveuse. Ce processus initie ensuite la libération présynaptique de calcium et la libération de neurotransmetteurs excitateurs sur les fibres nerveuses afférentes. Ces fibres transmettent ensuite des informations concernant l'ampleur du champ électrique détecté au système nerveux central du poisson.
Les ampoules intègrent des canaux potassiques activés par le calcium à grande conductance, appelés canaux BK. Les requins présentent une sensibilité exceptionnelle aux champs électriques, dépassant celle des poissons d'eau douce électrorécepteurs et de tous les autres animaux connus, avec un seuil de détection aussi bas que 5 nV/cm. L'hydrogel de collagène remplissant les canaux ampullaires possède l'une des capacités de conductivité protonique les plus élevées observées dans les matériaux biologiques. Composée de sulfate de kératane au sein d'une solution aqueuse à 97 %, cette substance présente une conductivité d'environ 1,8 mS/cm (0,18 S/m). Étant donné que tous les animaux génèrent un champ électrique par le biais de contractions musculaires, les poissons électrorécepteurs peuvent détecter de faibles stimuli électriques émanant de l'activité musculaire de leurs proies potentielles.
Les poissons-scies possèdent un plus grand nombre de pores ampullaires que tout autre poisson cartilagineux, ce qui en fait des spécialistes de l'électroréception. Les ampoules de Lorenzini chez le poisson-scie sont réparties sur toute la tête, sur les surfaces ventrale et dorsale de leur rostre s'étendant vers les branchies, et le long de la face dorsale de leur corps.
Magnétoréception
Les ampoules de Lorenzini facilitent également la réception des informations géomagnétiques. Compte tenu de la relation intrinsèque entre les champs magnétiques et électriques, la magnétoréception par induction électromagnétique dans les ampoules de Lorenzini est un mécanisme plausible. Une étude de 2004 a démontré cette capacité magnétoréceptive chez les poissons cartilagineux, comme en témoignent leurs réponses aux champs magnétiques générés artificiellement liés aux récompenses alimentaires. Environ huit requins étaient hébergés dans un réservoir doté d'un champ magnétique contrôlable. Au cours de la phase de conditionnement, le champ magnétique a été activé simultanément à la présentation des aliments à un emplacement cible spécifique sur le fond du réservoir. Par la suite, lors de la phase expérimentale, le champ a été activé sans apport de nourriture. Les observations ont révélé que les requins traversaient la zone cible de nourriture à une fréquence significativement plus élevée lorsque le champ magnétique était actif que lorsqu'il était éteint, indiquant ainsi un conditionnement réussi et leur capacité à détecter le champ magnétique.
La magnétoréception élucide potentiellement la capacité des requins et des raies à établir des routes migratoires précises et à connaître leur position géographique. Le noyau externe de la Terre produit un champ magnétique qui présente des fluctuations minimes tout au long de la vie de nombreuses espèces, mais qui varie toutefois nettement sur la surface de la planète, permettant aux animaux sensibles d'acquérir une compréhension générale de leur situation géographique. Les organismes possédant les ampoules de Lorenzini, entre autres, exploitent des composants tels que l'angle d'inclinaison, l'intensité du champ (force) et les magnitudes des champs magnétiques horizontaux et verticaux pour faire fonctionner une forme de système GPS interne. Ce système est crucial pour les espèces entreprenant de vastes migrations, comme les requins ; sans cela, leurs contributions aux écosystèmes naturels seraient considérablement réduites. En tant que prédateurs au sommet, ils régulent les populations de consommateurs primaires et leurs migrations facilitent le cycle et la redistribution des nutriments dans divers écosystèmes. L'absence de ce GPS interne pourrait conduire à l'abandon des emplacements d'origine des requins, perturbant ainsi l'équilibre de leurs réseaux trophiques respectifs.
Détection de la température
En 2003, il a été émis l'hypothèse que la substance semblable à du mucus à l'intérieur des tubes ampullaires pourrait fonctionner comme un semi-conducteur thermoélectrique, convertissant les variations de température en signaux électriques pour la détection des gradients thermiques par l'animal. Cependant, une étude de 2007 semble réfuter cette hypothèse. La question persistait et, d’ici 2023, les prédictions suggéraient que les ampoules de Lorenzini des requins pourraient détecter des différences de température aussi infimes que 0,001 Kelvin (un millième de degré). Un capteur artificiel fonctionnant sur un principe analogue démontre la capacité de détecter une différence de 0,01 Kelvin.
Le Knollenorgan est un type d'électrorécepteur non homologue identifié chez les poissons mormyridés.
- Knollenorgan – un type d'électrorécepteur non homologue, trouvé chez les poissons mormyridés